Every cell needs a way to turn stored genetic information into working molecules. DNA holds the instructions, but proteins do much of the cell’s daily labor: they form structures, speed up chemical reactions, move materials, send signals, and help regulate what happens next. Protein synthesis is the process that connects those two worlds. A gene’s DNA sequence is copied into messenger RNA, and that mRNA is then read by ribosomes to assemble a chain of amino acids.
The Central Dogma of Molecular Biology
The central dogma of molecular biology describes the usual flow of genetic information: DNA is copied into RNA, and RNA is used to build protein. The first step is transcription, where a cell makes an RNA copy of a gene. The second step is translation, where ribosomes read that RNA message and link amino acids in the correct order.
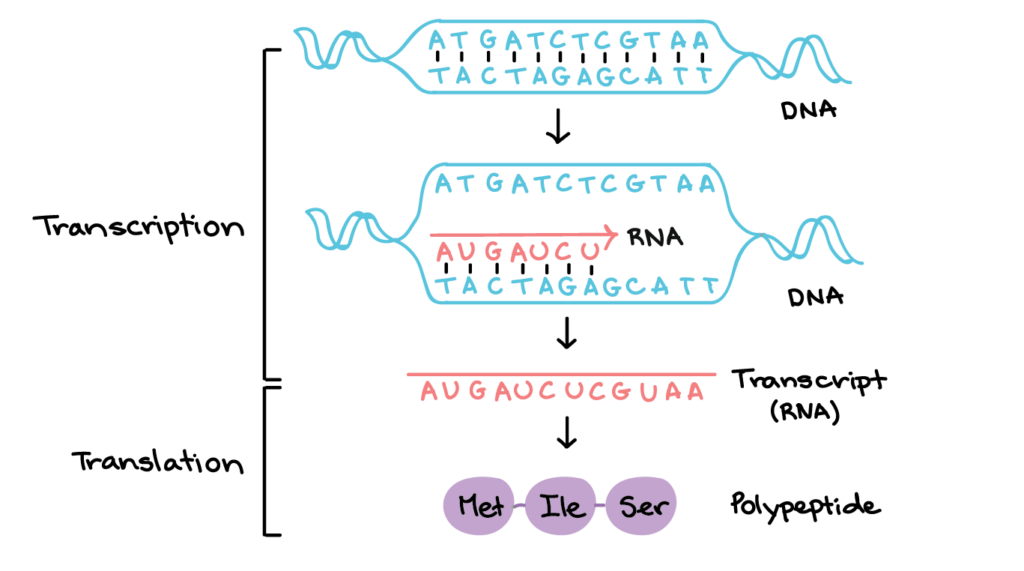
A gene is a stretch of DNA with information for a functional RNA or protein product. Not every gene becomes a protein, but protein-coding genes follow a clear route: the DNA sequence is transcribed into mRNA, the mRNA carries the message away from the DNA, and ribosomes translate that message into a polypeptide chain. The sequence matters because changing even one part of the message can sometimes change the amino acid sequence of the protein.
Transcription Copies DNA Into RNA
Transcription begins when RNA polymerase binds near a gene and uses one DNA strand as a template. DNA is double-stranded, but only one strand is copied for a particular gene. As RNA polymerase moves along the template strand, it builds a complementary RNA molecule using RNA nucleotides. In RNA, the base uracil pairs with adenine, while cytosine still pairs with guanine.
Initiation
In prokaryotes, RNA polymerase uses a sigma factor to recognize the promoter, a DNA region that signals where transcription should begin. After binding, RNA polymerase unwinds a small part of the DNA double helix so the template strand can be copied. Once the RNA chain is long enough, the sigma factor is released and transcription continues.
In eukaryotes, transcription is more heavily organized. General transcription factors bind near the promoter, often around a TATA box in genes that have one, and help position RNA polymerase. Together they form a transcription initiation complex. This careful setup helps the cell control which genes are active in different cell types and conditions.
Elongation
During elongation, RNA polymerase moves along the DNA template strand in the 3′ to 5′ direction. The growing RNA strand is made in the 5′ to 3′ direction as new ribonucleotides are added. RNA polymerase forms phosphodiester bonds between neighboring RNA nucleotides, creating a continuous strand. As the RNA strand lengthens, it peels away from the DNA, and the DNA helix rewinds behind the transcription bubble.
Termination
Transcription ends when RNA polymerase reaches a termination signal. In many prokaryotic genes, the RNA can fold into a hairpin structure that helps release the transcript from the polymerase. In eukaryotes, the RNA transcript is usually cut downstream of a polyadenylation signal, producing a pre-mRNA molecule that still needs processing before it can be translated.
RNA Processing Prepares the Message
In eukaryotic cells, the first RNA copy of a protein-coding gene is called pre-mRNA. Before it becomes mature mRNA, it is modified in several ways. These changes help protect the message, guide it out of the nucleus, and make it easier for ribosomes to recognize.
5′ Capping
A modified guanine nucleotide, called a 5′ cap, is added to the beginning of the pre-mRNA. The cap helps protect the RNA from breakdown, supports export from the nucleus, and plays a role when ribosomes begin translation. Without this kind of protection, many RNA messages would be degraded too quickly to be useful.
3′ Polyadenylation
A poly-A tail is added to the 3′ end of many eukaryotic mRNA molecules. This tail also improves stability and helps the mRNA move from the nucleus to the cytoplasm. A more stable mRNA can often be translated multiple times, allowing the cell to make many copies of a protein from one message.
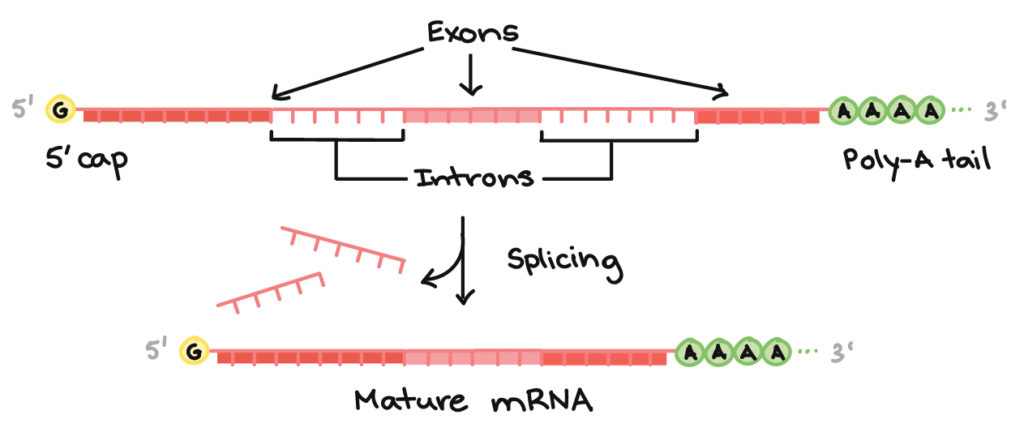
Splicing
Many eukaryotic genes contain introns and exons. Introns are removed from the pre-mRNA, while exons are joined together to make the mature message. This process is called splicing. In some genes, alternative splicing allows different combinations of exons to be joined, so one gene can help produce more than one protein version.
The Genetic Code Links Codons to Amino Acids
Translation depends on the genetic code. Ribosomes read mRNA in groups of three nucleotides called codons. Each codon corresponds to one amino acid or to a stop signal. Because there are four RNA bases, three-base codons create 64 possible combinations. That is enough to encode the 20 common amino acids used in proteins, plus start and stop signals.
The start codon is usually AUG, which codes for methionine and marks where translation begins. Three codons, UAA, UAG, and UGA, act as stop codons. The code is described as degenerate because more than one codon can specify the same amino acid. It is also unambiguous because a particular codon normally specifies only one amino acid or signal.
Translation Builds the Protein Chain
Translation happens at ribosomes. The ribosome reads the mRNA sequence while transfer RNA molecules, or tRNAs, bring the matching amino acids. Each tRNA has an anticodon that pairs with an mRNA codon and carries a specific amino acid. As codons are read in order, the ribosome links amino acids together to form a growing polypeptide chain.
Before translation can proceed efficiently, amino acids are attached to their correct tRNAs. This step is called amino acid activation, and it helps make sure the right amino acid is delivered for each codon. Accuracy matters: the sequence of amino acids determines how the protein folds and what it can do.
Initiation
In prokaryotes, the small ribosomal subunit binds near a sequence called the Shine-Dalgarno sequence, which helps position the ribosome near the start codon. The initiator tRNA carries a modified form of methionine and pairs with the start codon. The large ribosomal subunit then joins, forming a complete initiation complex.
In eukaryotes, the small ribosomal subunit usually binds near the 5′ cap and scans along the mRNA until it reaches an appropriate start codon. The initiator tRNA carries methionine. Once the start codon is recognized, the large ribosomal subunit attaches and translation can begin.
Elongation
During elongation, a tRNA carrying the next amino acid enters the ribosome and pairs with the next mRNA codon. The ribosome forms a peptide bond between the growing chain and the new amino acid. Then the ribosome shifts forward by one codon, the empty tRNA exits, and another tRNA can enter. This cycle repeats until the full coding sequence has been read.
Termination
Translation ends when the ribosome reaches a stop codon. No tRNA carries an amino acid for a stop codon. Instead, a release factor binds, the completed polypeptide is released, and the ribosomal subunits separate from the mRNA. The protein chain can then begin folding into the shape it needs to work properly.
What Happens After Translation
A newly made polypeptide is not always ready to function immediately. Many proteins must fold into precise shapes, sometimes with help from other proteins called chaperones. Some proteins are chemically modified after translation, which can change where they go in the cell, how active they are, or how long they last.
Cells also need ways to remove proteins that are damaged, misfolded, or no longer needed. In eukaryotes, one major pathway marks proteins with a small protein called ubiquitin. Tagged proteins can then be broken down by proteasomes, allowing the cell to recycle their amino acids and keep protein levels under control.
Protein synthesis is powerful because it turns a sequence of bases into a molecule that can actually do work. Transcription preserves the information in a portable RNA message. Translation reads that message in codons and builds a chain of amino acids. From there, folding and modification turn the chain into a functional protein, connecting genetic instructions to the living cell’s structure and activity.